
Head

Prof. Dr. Pamela Fischer-Posovszky
Ulm University Medical Center
Department of Pediatrics and Adolescent Medicine
Eythstr. 24 (Research Lab, House 16)
89075 Ulm, Germany
phone: +49-731-500 57415
e-mail: pamela.fischer@uniklinik-ulm.de

| Name | Function | |
|---|---|---|
| Prof. Dr. Pamela Fischer-Posovszky | Head | pamela.fischer@uniklinik-ulm.de |
| Dr. Julia Zinngrebe | Deputy Head | julia.zinngrebe@uniklinik-ulm.de |
| Johanna Gernoth | Student | johanna.gernoth@uniklinik-ulm.de |
| Alexandra Killian | TA | alexandra.killian@uniklinik-ulm.de |
| Dr. Paul Knoop | Post-Doc | paul.knoop@uniklinik-ulm.de |
| Hanna Meid | Phd Student | hanna.meid@uniklinik-ulm.de |
| Corinna Stiegeler | Student | corinna.stiegeler@uniklinik-ulm.de |
| Dr. Ulrich Stifel | Post-Doc | ulrich.stifel@uniklinik-ulm.de |
| Prof. Dr. Daniel Tews | Deputy Head Core Facility | daniel.tews@uniklinik-ulm.de |
| Anja Werberger | Phd Student | anja.werberger@uniklinik-ulm.de |
| Niklas Widmann | Student | niklas.widmann@uniklinik-ulm.de |
News & Events

Mit diesem Forschungspreis zeichnet die Universität Ulm einmal jährlich exzellente Nachwuchswissenschaftlerinnen aus, die durch ihre herausragende wissenschaftliche Arbeit die Universität Ulm als Wissenschaftsinstitution mitgestalten und eine bedeutende Vorbildfunktion für andere Wissenschaftlerinnen sowie Studentinnen übernehmen. Wir gratulieren!



Paul and Pam participate in the kick-off meeting of the DZKJ at the town hall.
For more info please see:
www.uni-ulm.de/home/uni-aktuell/article/ulmer-dzkj-standort-stellt-sich-der-oeffentlichkeit-vor/

Research Profile
Our laboratory explores how adipose tissue develops and functions in health and disease.
In our early work, we focused on fundamental questions such as: What controls the number of fat cells? Which genes determine the amount of body fat? And how do different types of fat—white, brown, and beige—develop in humans?
More recently, our research has shifted toward understanding how adipose tissue communicates with the rest of the body. We study its role in metabolism and inflammation, as well as its interactions with other organs. Current projects examine:
• the connection between adipose tissue and the brain,
• the role of adipose tissue during trauma,
• how small regulatory molecules (microRNAs) influence adipose tissue,
• how linear ubiquitin affects adipocyte function,
• the presence and role of adipocytes in bone marrow, and
• the effects of excess glucocorticoids on adipose tissue
Beyond adipose biology, we are also investigating the metabolism of immune cells and how this might be linked to mental health.
In addition, our group operates the Core Facility “Extracellular Flux Analyzer”, which provides advanced technology to measure cellular metabolism.

The adipose tissue–brain axis represents a critical line of communication between peripheral metabolism and central regulation of energy balance, stress responses, and immune function. Our lab investigates how adipose tissue signals - particularly hormones and cytokines - are sensed and processed by the brain, and how central responses in turn influence adipose tissue behavior.
Discovery of novel treatment options for obesity
We explore this bidirectional communication under both physiological and pathological conditions, including trauma, metabolic stress, and inflammation. By integrating neuroendocrinology, immunology, and metabolism, we aim to reveal how disruptions in the adipose–brain axis contributes to systemic dysregulation. Our goal is to identify novel mechanisms through which peripheral metabolic organs influence central nervous system functions, with implications for treating metabolic and inflammatory diseases.
Leptin
One crucial player in the adipose tissue-brain axis is leptin. The hormone is produced by adipocytes to signal the energy state of the body to the brain. Congenital leptin deficiency is caused by variants in the leptin gene resulting in different defects in protein expression, secretion, or function. Affected patients suffer from hyperphagia and severe, early-onset obesity (Hinney et al., Nat Rev Endocrinol 2022).
Together with physicians at the Division of Pediatric Endocrinology and Diabetes and other collaborators, we characterized new forms of leptin deficiency. Patients with functional leptin deficiency have high circulating leptin levels, but the variant hormone is biologically inactive as it does not bind to ist receptor. Therefore, it is not capable of delivering a satiety signal to the brain. The disease can be treated by daily injections of human recombinant leptin leading to a normalization of eating behavior and rapid weight loss (Wabitsch et al., New Engl J Med 2015).
Patients with an antagonistic leptin hormone have high circulating leptin levels. The variant hormone can bind to but does not activate the leptin receptor. Therefore, these variants act as competitive antagonists in the presence of nonvariant leptin, which complicates treatment (Funcke et al., New Engl J Med 2023).
We aim to elucidate novel biological functions of leptin and its signaling properties in the context of obesity and immunology.
Funding:
Boehringer Ingelheim Ulm University BioCenter (BIU 3.0)
Ulm University, Medical Faculty (Bausteinprogramm)
Publications:
Classification of Congenital Leptin Deficiency.
von Schnurbein J, Zorn S, Nunziata A, Brandt S, Moepps B, Funcke JB, Hussain K, Farooqi IS, Fischer-Posovszky P, Wabitsch M. J Clin Endocrinol Metab. 2024 Sep 16;109(10):2602-2616. doi: 10.1210/clinem/dgae149.PMID: 38470203
Rare Antagonistic Leptin Variants and Severe, Early-Onset Obesity.
Funcke JB#, Moepps B#, Roos J#, von Schnurbein J, Verstraete K, Fröhlich-Reiterer E, Kohlsdorf K, Nunziata A, Brandt S, Tsirigotaki A, Dansercoer A, Suppan E, Haris B, Debatin KM, Savvides SN, Farooqi IS, Hussain K, Gierschik P, Fischer-Posovszky P*, Wabitsch M* (#,* equal contribution) N Engl J Med. 2023 Jun 15;388(24):2253-2261. doi: 10.1056/NEJMoa2204041.PMID: 37314706
The promise of new anti-obesity therapies arising from knowledge of genetic obesity traits.
Hinney A, Körner A, Fischer-Posovszky P. Nat Rev Endocrinol. 2022 Oct;18(10):623-637. doi: 10.1038/s41574-022-00716-0.
Early childhood BMI trajectories in monogenic obesity due to leptin, leptinreceptor, and melanocortin 4 receptor deficiency.
Kohlsdorf K, Nunziata A, Funcke JB, Brandt S, von Schnurbein J, Vollbach H, Lennerz B, Fritsch M, Greber-Platzer S, Fröhlich-Reiterer E, Luedeke M, Borck G, Debatin KM, Fischer-Posovszky P, Wabitsch M. Int J Obes (Lond). 2018 Sep;42(9):1602-1609. doi: 10.1038/s41366-018-0049-6.
Biologically inactive leptin and early-onset extreme obesity.
Wabitsch M, Funcke JB, Lennerz B, Kuhnle-Krahl U, Lahr G, Debatin KM, Vatter P, Gierschik P, Moepps B, Fischer-Posovszky P. N Engl J Med. 2015 Jan 1;372(1):48-54. doi: 10.1056/NEJMoa1406653.




Adipose tissue plays a pivotal role in the systemic response to trauma (Wrba et al., Cell Mol Life Sci, 2022). In our lab, we investigate how thorax trauma and hemorrhagic shock influence adipose tissue biology and function. These severe injuries trigger complex inflammatory and metabolic cascades, in which adipose tissue emerges not merely as a passive target but as an active responder and regulator. We focus on how trauma-induced signals reprogram adipocytes and their interactions with immune and stromal cells. Our research aims to define how these changes contribute to systemic inflammation, immune dysregulation, and organ dysfunction following trauma (Roos et al., Cell Death Dis, 2024). Using in-vivo trauma models, molecular profiling, and metabolic analysis, we seek to uncover adipose-specific mechanisms that influence patient outcomes and may serve as therapeutic targets in trauma care.
Funding:
CRC1149, project A09
Principal investigators:
Jun.Prof. Dr. Rebecca Halbgebauer, Institute of Clinical and Experimental Trauma Immunology & Prof. Dr. Pamela Fischer-Posovszky
www.uni-ulm.de/einrichtungen/sfb-1149/
Publications:
Trauma-associated extracellular histones mediate inflammation via a MYD88-IRAK1-ERK signaling axis and induce lytic cell death in human adipocytes.
Roos J, Zinngrebe J, Huber-Lang M, Lupu L, Schmidt MA, Strobel H, Westhoff MA, Stifel U, Gebhard F, Wabitsch M, Mollnes TE, Debatin KM, Halbgebauer R, Fischer-Posovszky P.
Cell Death Dis. 2024 Apr 23;15(4):285. doi: 10.1038/s41419-024-06676-9.
Adipose tissue: a neglected organ in the response to severe trauma?
Wrba L, Halbgebauer R, Roos J, Huber-Lang M, Fischer-Posovszky P.
Cell Mol Life Sci. 2022 Mar 26;79(4):207. doi: 10.1007/s00018-022-04234-0.

MicroRNAs (miRNAs) are short, non-coding RNA molecules, typically 18–25 nucleotides in length. In recognition of their groundbreaking discovery of miRNAs in 1993 and the elucidation of their role in post-transcriptional gene regulation, Victor Ambros and Gary Ruvkun were awarded the Nobel Prize in 2024. Since their initial discovery, miRNAs have been shown to play key roles in the regulation of gene expression, influencing critical cellular processes such as differentiation, proliferation, and survival. Our research has contributed to advancing our understanding on the function of miRNAs in adipose tissue (Roos et al., Sci Rep, 2016; Wu H et al., Cells, 2021; Roos J et al., Cell Mol Life Sci, 2021). We are currently expanding this work by investigating the involvement of miRNAs in adipocyte metabolism and exploring their potential as innovative therapeutic targets in the context of obesity.
Funding:
Boehringer Ingelheim Ulm University BioCenter (BIU 2.0)
Publications:
microRNA-27a-3p but Not -5p Is a Crucial Mediator of Human Adipogenesis.
Wu H, Pula T, Tews D, Amri EZ, Debatin KM, Wabitsch M, Fischer-Posovszky P, Roos J.
Cells. 2021 Nov 17;10(11):3205. doi: 10.3390/cells10113205.
miR-146a regulates insulin sensitivity via NPR3.
Roos J, Dahlhaus M, Funcke JB, Kustermann M, Strauss G, Halbgebauer D, Boldrin E, Holzmann K, Möller P, Trojanowski BM, Baumann B, Debatin KM, Wabitsch M, Fischer-Posovszky P.
Cell Mol Life Sci. 2021 Mar;78(6):2987-3003. doi: 10.1007/s00018-020-03699-1.
Human adipocyte differentiation and composition of disease-relevant lipids are regulated by miR-221-3p.
Ahonen MA, Asghar MY, Parviainen SJ, Liebisch G, Höring M, Leidenius M, Fischer-Posovszky P, Wabitsch M, Mikkola TS, Törnquist K, Savolainen-Peltonen H, Haridas PAN, Olkkonen VM.
Biochim Biophys Acta Mol Cell Biol Lipids. 2021 Jan;1866(1):158841. doi: 10.1016/j.bbalip.2020.158841.
miR-107 inhibits CDK6 expression, differentiation, and lipid storage in human adipocytes.
Ahonen MA, Haridas PAN, Mysore R, Wabitsch M, Fischer-Posovszky P, Olkkonen VM.
Mol Cell Endocrinol. 2019 Jan 5;479:110-116. doi: 10.1016/j.mce.2018.09.007.
A computational biology approach of a genome-wide screen connected miRNAs to obesity and type 2 diabetes.
Gottmann P, Ouni M, Saussenthaler S, Roos J, Stirm L, Jähnert M, Kamitz A, Hallahan N, Jonas W, Fritsche A, Häring HU, Staiger H, Blüher M, Fischer-Posovszky P, Vogel H, Schürmann A.
Mol Metab. 2018 May;11:145-159. doi: 10.1016/j.molmet.2018.03.005.
Circulating levels of miR-122 and nonalcoholic fatty liver disease in pre-pubertal obese children.
Brandt S, Roos J, Inzaghi E, Kotnik P, Kovac J, Battelino T, Cianfarani S, Nobili V, Colajacomo M, Kratzer W, Denzer C, Fischer-Posovszky P, Wabitsch M.
Pediatr Obes. 2018 Mar;13(3):175-182. doi: 10.1111/ijpo.12261.
miR-146a-mediated suppression of the inflammatory response in human adipocytes.
Roos J, Enlund E, Funcke JB, Tews D, Holzmann K, Debatin KM, Wabitsch M, Fischer-Posovszky P.
Sci Rep. 2016 Dec 6;6:38339. doi: 10.1038/srep38339.
Functional Significance and Predictive Value of MicroRNAs in Pediatric Obesity: Tiny Molecules with Huge Impact?
Fischer-Posovszky P, Roos J, Kotnik P, Battelino T, Inzaghi E, Nobili V, Cianfarani S, Wabitsch M.
Horm Res Paediatr. 2016;86(1):3-10. doi: 10.1159/000444677.

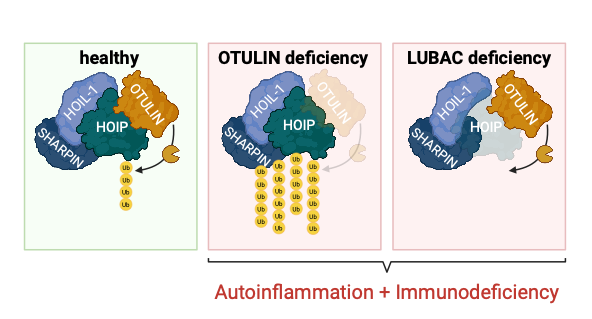
Linear ubiquitin chains play a crucial role in regulating inflammatory processes in the human body (Zinngrebe J. et al., EMBO Reports, 2014). These chains are assembled by the Linear UBiquitin chain Assembly Complex (LUBAC), which consists of SHARPIN, HOIL-1, and HOIP, and are specifically removed by the deubiquitinase OTULIN (OTU Deubiquitinase with LINear linkage specificity). Dysregulation of linear ubiquitination – e.g., due to mutations in LUBAC or OTULIN – can lead to severe autoinflammation and/or immunodeficiency in affected patients (Zinngrebe J. et al., EMBO Mol Med, 2022). Although adipose tissue is frequently affected, the underlying molecular mechanisms remain poorly understood. Gaining deeper insights into these processes may open up new therapeutic avenues for affected patients.
Funding:
Deutsche Forschungsgemeinschaft
Publications:
Linear ubiquitination prevents lipodystrophy and obesity-associated metabolic syndrome.
Hildebrandt X, Veli Ö, Hyoubi A, Zinngrebe J, Abdallah AT, Rodefeld J, Hoffmann A, Gardeweg L, Kaya Ö, Wagner E, Lindhorst A, Poggenberg M, Wang Y, Dimmler J, Schillings J, Koci P, Bonechi F, Capuccino LV, Kiefer C, Kelepouras K, Ghosh A, Noé F, Wolfrum C, Singer M, Liccardi G, Luedde T, Yavas A, Ghallab A, Hengsler JG, Antczak P, Gericke M, Winkels H, Blüher M, Walczak H, Annibaldi A, Fischer-Posovszky P, Peltzer N.
Sci Adv. 2025 Sep 19;11(38):eadw2539. doi: 10.1126/sciadv.adw2539.
Compound heterozygous variants in OTULIN are associated with fulminant atypical late-onset ORAS.
Zinngrebe J, Moepps B, Monecke T, Gierschik P, Schlichtig F, Barth TFE, Strauß G, Boldrin E, Posovszky C, Schulz A, Beringer O, Rieser E, Jacobsen EM, Lorenz MR, Schwarz K, Pannicke U, Walczak H, Niessing D, Schuetz C, Fischer-Posovszky P, Debatin KM.
EMBO Mol Med. 2022 Mar 7;14(3):e14901. doi: 10.15252/emmm.202114901
TLRs Go Linear - On the Ubiquitin Edge.
Zinngrebe J, Walczak H.
Trends Mol Med. 2017 Apr;23(4):296-309. doi: 10.1016/j.molmed.2017.02.003.
--LUBAC deficiency perturbs TLR3 signaling to cause immunodeficiency and autoinflammation.
Zinngrebe J, Rieser E, Taraborrelli L, Peltzer N, Hartwig T, Ren H, Kovács I, Endres C, Draber P, Darding M, von Karstedt S, Lemke J, Dome B, Bergmann M, Ferguson BJ, Walczak H.
J Exp Med. 2016 Nov 14;213(12):2671-2689. doi: 10.1084/jem.20160041.
Ubiquitin in the immune system.
Zinngrebe J, Montinaro A, Peltzer N, Walczak H.
EMBO Rep. 2014 Jan;15(1):28-45. doi: 10.1002/embr.201338025.


Bone marrow adipocytes (BMAd), once believed to lack specific function, are increasingly recognized as important regulators of hematopoiesis (Zinngrebe J. et al., Leukemia, 2020). Today, we know that BMAd - similar to adipocytes in white adipose tissue depots - store energy and secrete important signaling molecules. BMAd exhibit remarkable plasticity and form in response to various external stimuli, such as steroids, obesity, or during aging and under anti-leukemic therapy. The intricate interplay between BMAd and other cells in the bone marrow microenvironment remains incompletely understood. Therefore, we study BMAd under different physiological and pathological conditions by state-of-the-art sequencing techniques to elucidate previously unknown interaction routes between adipocytes and cells of the hematopoietic system.
Funding:
Startup funding by CRC1074, CRC1506
Publications:
Bridging the marrow: a co-culture-platform of leukemia cells and MS5-derived stromal cells or adipocytes.
Zinngrebe J, Brenner ED, Schlichtig F, Stifel U, Tews D, Falk J, Schlotter D, Fitzel R, Meyer LH, Debatin KM, Fischer-Posovszky P.
Cell Death Discov. 2025 Aug 5;11(1):366. doi: 10.1038/s41420-025-02631-5.
Adipocytes in hematopoiesis and acute leukemia: friends, enemies, or innocent bystanders?
Zinngrebe J, Debatin KM, Fischer-Posovszky P.
Leukemia. 2020 Sep;34(9):2305-2316. doi: 10.1038/s41375-020-0886-x.
Cushing’s syndrome (CS) is a rare endocrine disorder characterized by chronic exposure to excess glucocorticoids (GC), leading to a constellation of clinical features including central obesity, insulin resistance, muscle wasting, and hypertension. However, the transient development of CS can also be observed under external glucocorticoid treatment. Despite the prominence of abnormal fat distribution—particularly visceral adiposity—in CS, the underlying mechanisms driving these changes in adipose tissue remain incompletely understood. Our research group aims to integrate clinical observations and material with basic and translational research to explore how glucocorticoid excess alters adipose tissue structure, function, and endocrine signaling. Key areas of interest include depot-specific differences, the role of adipocyte hypertrophy and hyperplasia, inflammatory cell infiltration and changes in preadipocytes.
Our multidisciplinary approach leverages advanced imaging techniques, transcriptomic and proteomic and metabolomics profiling, and patient-derived adipose samples. To achieve this, we have multiple collaborations across institutions to facilitate the sharing of clinical data and biobank resources as well as technologies.
The long-term goal of the working group is to clarify the contribution of adipose tissue to the systemic metabolic dysfunction seen in CS and to understand the long-lasting metabolic effects of chronic GC exposure on adipose tissue.
Funding:
Deutsche Gesellschaft für Endokrinologie
Publications:
Metabolic rewiring promotes anti-inflammatory effects of glucocorticoids.
Auger JP, Zimmermann M, Faas M, Stifel U, Chambers D, Krishnacoumar B, Taudte RV, Grund C, Erdmann G, Scholtysek C, Uderhardt S, Ben Brahim O, Pascual Maté M, Stoll C, Böttcher M, Palumbo-Zerr K, Mangan MSJ, Dzamukova M, Kieler M, Hofmann M, Blüml S, Schabbauer G, Mougiakakos D, Sonnewald U, Hartmann F, Simon D, Kleyer A, Grüneboom A, Finotto S, Latz E, Hofmann J, Schett G, Tuckermann J, Krönke G.
Nature. 2024 May;629(8010):184-192. doi: 10.1038/s41586-024-07282-7.
Unique Gene Expression Signature in Periadrenal Adipose Tissue Identifies a High Blood Pressure Group in Patients With Cushing Syndrome.
Stifel U*, Vogel F*, Caratti G, Reincke M, Tuckermann J.
Hypertension. 2023 Nov;80(11):2333-2344. doi: 10.1161/HYPERTENSIONAHA.123.21185.
Glucocorticoid activation of anti-inflammatory macrophages protects against insulin resistance.
Caratti G, Stifel U, Caratti B, Jamil AJM, Chung KJ, Kiehntopf M, Gräler MH, Blüher M, Rauch A, Tuckermann JP.
Nat Commun. 2023 Apr 20;14(1):2271. doi: 10.1038/s41467-023-37831-z.
Macrophagic AMPKα1 orchestrates regenerative inflammation induced by glucocorticoids.
Caratti G, Desgeorges T, Juban G, Stifel U, Fessard A, Koenen M, Caratti B, Théret M, Skurk C, Chazaud B, Tuckermann JP, Mounier R.
EMBO Rep. 2023 Feb 6;24(2):e55363. doi: 10.15252/embr.202255363.
Novel insights into the regulation of cellular catabolic metabolism in macrophages through nuclear receptors.
Stifel U, Caratti G, Tuckermann J.
FEBS Lett. 2022 Oct;596(20):2617-2629. doi: 10.1002/1873-3468.14474.
A macrophage-hepatocyte glucocorticoid receptor axis coordinates fasting ketogenesis.
Loft A, Schmidt SF, Caratti G, Stifel U, Havelund J, Sekar R, Kwon Y, Sulaj A, Chow KK, Alfaro AJ, Schwarzmayr T, Rittig N, Svart M, Tsokanos FF, Maida A, Blutke A, Feuchtinger A, Møller N, Blüher M, Nawroth P, Szendrödi J, Færgeman NJ, Zeigerer A, Tuckermann J, Herzig S.
Cell Metab. 2022 Mar 1;34(3):473-486.e9. doi: 10.1016/j.cmet.2022.01.004.
Glucocorticoids coordinate macrophage metabolism through the regulation of the tricarboxylic acid cycle.
Stifel U, Wolfschmitt EM, Vogt J, Wachter U, Vettorazzi S, Tews D, Hogg M, Zink F, Koll NM, Winning S, Mounier R, Chazaud B, Radermacher P, Fischer-Posovszky P, Caratti G, Tuckermann J.
Mol Metab. 2022 Mar;57:101424. doi: 10.1016/j.molmet.2021.101424.
Imbalances during the critical early developmental stages of children can have significant repercussions on their health. At the newly established German Center for Pediatric and Adolescent Health (DZKJ), our research group is dedicated to investigating inborn errors of immunity and elucidating the alterations in the immune system from early childhood through adolescence. Understanding of normal immune function, immune deficiency and immune regulation is key for a range of common diseases with a strong socio-economic impact.
Increasing evidence demonstrates mental disease to be programmed during childhood through imprinting of psycho-social stress within biomolecular processes including immune-metabolic processes and mitochondrial changes. Within the DZKJ, we are examining the interconnections between mental health, metabolism, and immunological factors.
Utilizing methodologies such as single-cell metabolic flux analysis, real-time respirometry, and -omics approaches, we aim to identify the specific cell types responsible for the observed imbalances in mental health and immunity. Ultimately, our goal is to define the endotypes of these complex conditions and develop novel therapeutics to advance personalized medicine.
Funding:
German Center for Child an Adolescent Health (DZKJ), Federal Ministry of Research, Technology and Space (Bundesministrerium für Forschung, Technologie und Raumfahrt, BMFTR)


The ULMTeC core facility Extracellular Flux Analyzer offers support for real-time analysis of energy metabolism of live cells. Using a “Seahorse” XFe96 analyzer, automatic measurements of oxygen consumption rate (OCR), proton efflux rate (PER) or extracellular acidification rate (ECAR), as well as ATP production rates can be performed by label-free detection, interrogating key cellular functions like mitochondrial respiration and glycolysis. Measurements provide a systems-level view of cellular metabolic function relevant in research areas involving metabolism, oncology, neurology, immunology, and many more.
Within the core facility, we provide technical as well as scientific counselling regarding design, implementation and evaluation of experiments. If desired, experiments can also be carried out as full service entirely by the core facility.
Detailed information can be found here:
www.uni-ulm.de/med/ulmtec/cf/flux/
